Cours : La complexification des génomes
Mise à jour 04/01/2026 par JBoscq
Introduction :
La méiose et la fécondation permettent de brasser les gènes au sein d’une espèce créant de la diversité entre les individus. Si ce brassage est un atout en terme d’adaptation de l’espèce, de nouvelles mutations transmissibles peuvent apparaître créant de nouvelles populations, voire de nouvelles espèces, à condition que ces dernières confèrent un avantage et non l’inverse. Mais l’énorme diversité des êtres vivants ne peut s’expliquer uniquement par ces modifications. Il doit donc exister d’autres mécanismes permettant de complexifier le génome des êtres vivants : c’est ce que nous allons voir à travers les transferts horizontaux et les endosymbioses.
Photo de couverture : bactériophages d’après futurascience
Problème : Comment complexifier le genome d’une espèce, permettant une meilleure adaptation à l’environnement, à partir de nouveaux éléments génétiques ?
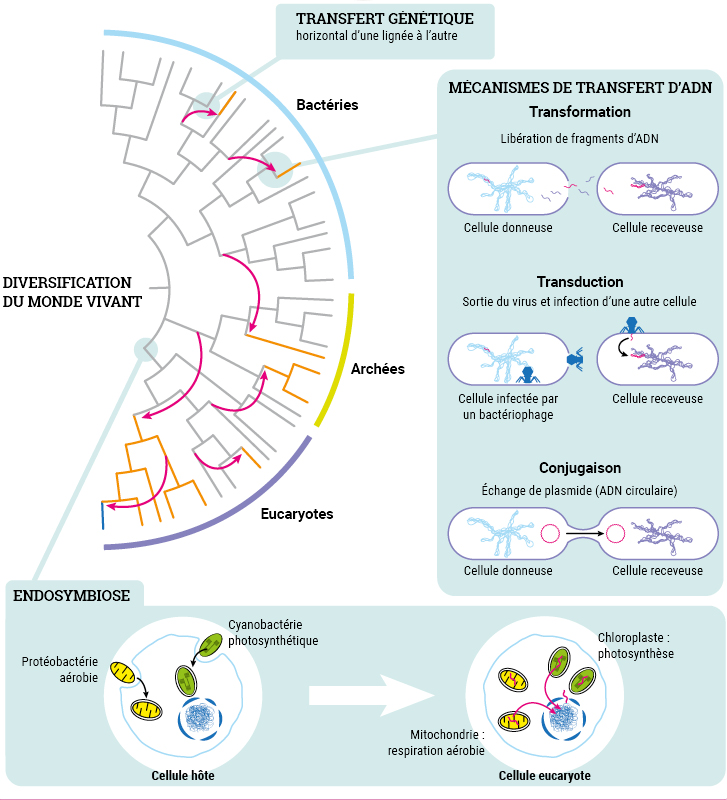
I- Les transferts horizontaux
A- Universalité structurale et fonctionnelle de l’ADN
La molécule d’ADN est le support de l’information génétique des cellules. Elle présente une organisation et un fonctionnement universels au sein du monde vivant. C’est d’ailleurs l’un des arguments qui permet d’envisager une origine commune de tous les êtres vivants. Ainsi, l’information génétique présente la même structure, et le même codage de l’information chez tous les êtres vivants, que l’on soit une bactérie ou un éléphant. Ce qui va changer d’un individu à un autre, c’est une différence d’allèles. Ce qui va changer d’une espèce à une autre, c’est aussi la qualité des gènes, mais également la quantité de gènes exprimés.
C’est grâce à l’universalité de l’ADN que nous pouvons réaliser des transgenèses en biotechnologie.
Doc. 1 : Universalité de la molécule d’ADN.
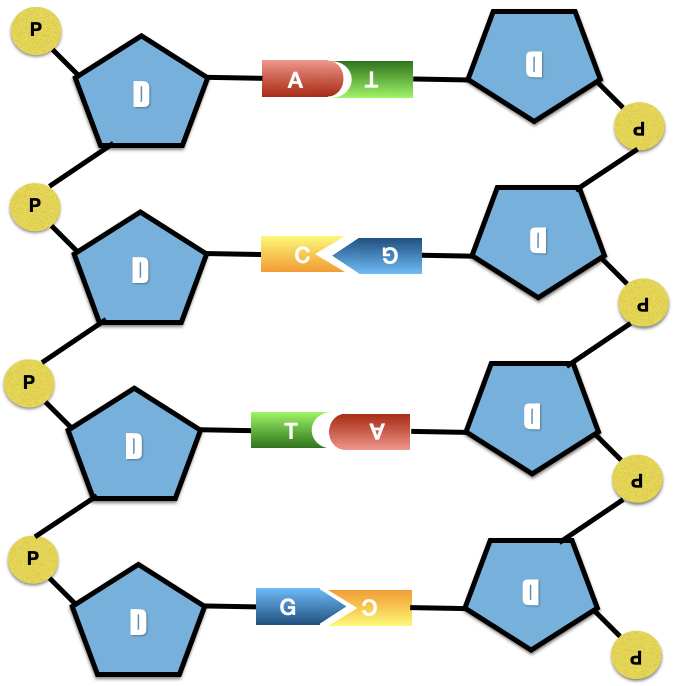
D’après JBoscq
B- La découverte de la transformation bactérienne

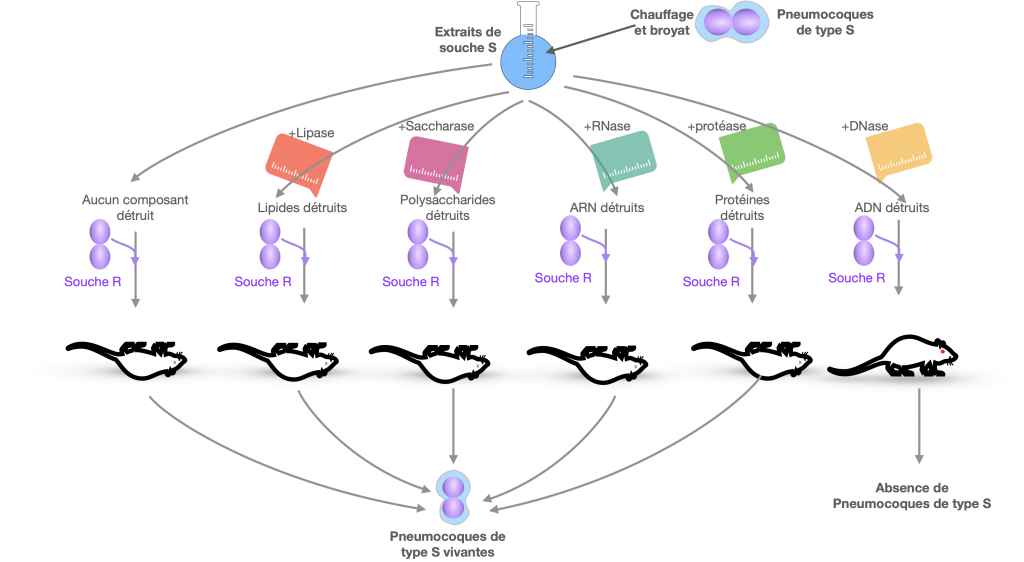
Expériences de Griffith

Frederick Griffith fit une observation inattendue au cours d‘expériences réalisées en 1928 sur des bactéries de pneumocoques (Streptococcus pneumoniae). Cette bactérie est responsable de la pneumonie chez l’Etre Humain, mais tue également les souris. Griffith découvrit une souche moins virulente (R = rugueuse) avec laquelle il réalisa des séries d’expériences ainsi qu’avec la souche virulente (S = lisse). Griffith montra qu’il existe un principe transformant pouvant passer d’une population de bactéries virulente à une autre non virulente et qui à son tour devient virulente. Autrement dit la souche R se transforme en souche S.
L’étape suivante consistait à identifier le support moléculaire responsable de cette transformation. Cela fut résolu en 1944 par Oswald Avery et 2 de ses collaborateurs, Colin McLeod et Maclyn McCarthy qui ont repris les expériences de Griffith et ont montré parmi différentes molécules, que seul l’ADN était responsable de cette propriété transformante.
L’autre enseignement est que les bactéries sont capables de s’échanger de l’ADN avec leur milieu.
Doc. 2 : Expériences de Avery, MacLeod et Mac Carthy – 1944
C- Définitions et mécanismes des transferts horizontaux

Définitions :
Transfert vertical : Dans le cadre d’une reproduction sexuée, le brassage des gènes s’effectue et nous héritons des gènes de nos parents. Ce transfert de gènes est qualifié de transfert vertical car il fait descendre les gènes de génération en génération.
Transfert horizontal : Le transfert horizontal de gènes est un processus qui permet à un organisme d’intégrer du matériel génétique provenant d’un autre organisme sans en être apparenté. C’est un mécanisme très fréquent entre différentes bactéries d’espèces différentes, mais cela existe aussi entre eucaryotes unicellulaires et entre les virus et tous les organismes vivants.
Les mécanismes de transferts horizontaux sont fréquents et multiples, notamment entre procaryotes. Les gènes assimilés par les organismes leur confèrent de nouveaux phénotypes. Cela influe alors sur les populations et leur adaptation à l’environnement.
Chez des bactéries d’espèces différentes, on peut distinguer 3 mécanismes principaux de transferts horizontaux.
Doc. 3 : Schéma d’une bactérie
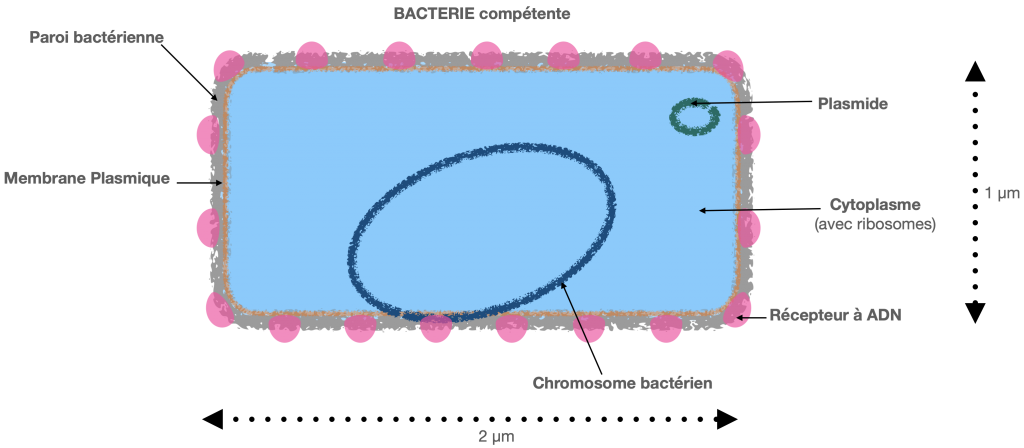
1- la transformation.
C’est le mécanisme découvert par Griffith en 1928. Il s’agit d’un transfert passif d’ADN d’une bactérie donatrice à une bactérie réceptrice, dite en état de compétence. Le transfert est partiel, autrement dit, l’ADN assimilé n’est pas l’ADN complet de la bactérie morte. L’ADN issu de la bactérie morte se fixe sur la paroi de la bactérie qui assimile cet ADN, au niveau de sites récepteurs et dans des conditions strictes de l’environnement.
Malgré tout, ce transfert horizontal, joue un rôle non négligeable dans l’assimilation ou l’apprentissage de la résistance aux antibiotiques des pneumocoques.
Doc. 4 : Schématisation de la transformation.
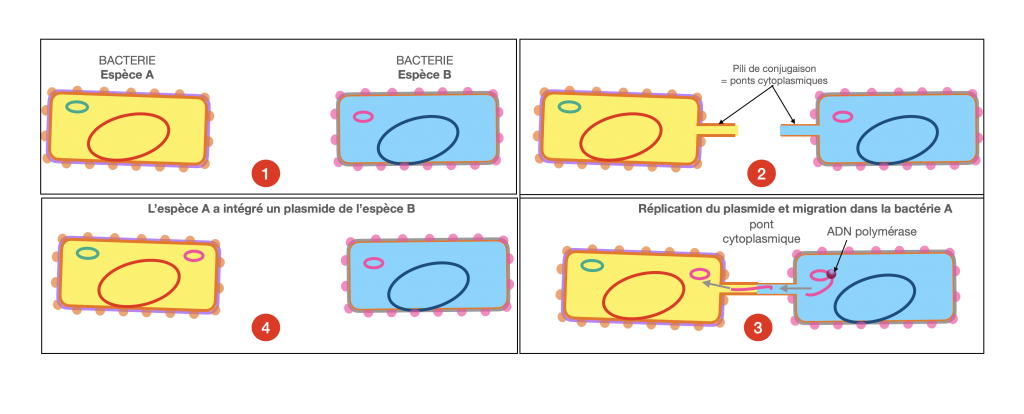
2- la conjugaison
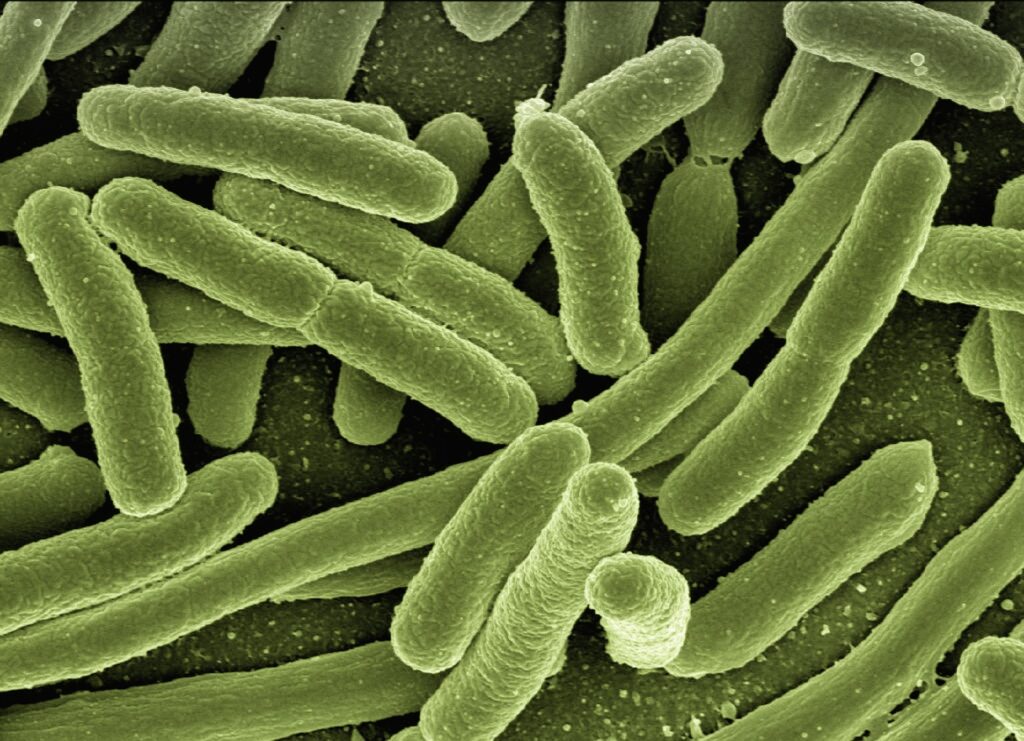
La conjugaison est un mode de communication entre bactéries ou plutôt un mode d’échange de matériels génétiques mobiles qu’on appelle plasmides. Un pont cytoplasmique se crée entre les 2 bactéries tandis qu’un plasmide a été répliqué. La copie du plasmide passe par le pont cytoplasmique d’une bactérie à l’autre. Il s’agit du principal facteur d’évolution des bactéries, en particulier pour l’acquisition de la résistance aux antibiotiques.
Définition :
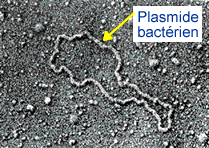
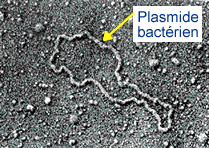
Plasmide: Les plasmides sont des petits fragments d’ADN circulaire présents dans la cellule bactérienne et indépendants du génome bactérien.
Complément :
Les plasmides présentent les caractéristiques suivantes :
- ➡Leur ADN est bicaténaire, circulaire avec un nombre de nucléotides inférieur à 10 kb (1 kilobase = kb = 1000 nucléotides).
{kind=link}
- ➡Le nombre de plasmides dans une cellule bactérienne peut être considérable (plusieurs centaines).
- ➡Les plasmides portent normalement des gènes qui leur confèrent un avantage sélectif, par exemple une résistance à un antibiotique.
- ➡Leur réplication est indépendante de celle du génome bactérien.
Doc. 5 : Schématisation de la conjugaison.
Doc. 6: photos obtenues en Microscopie électronique de conjugaisons bactériennes chez E.coli.
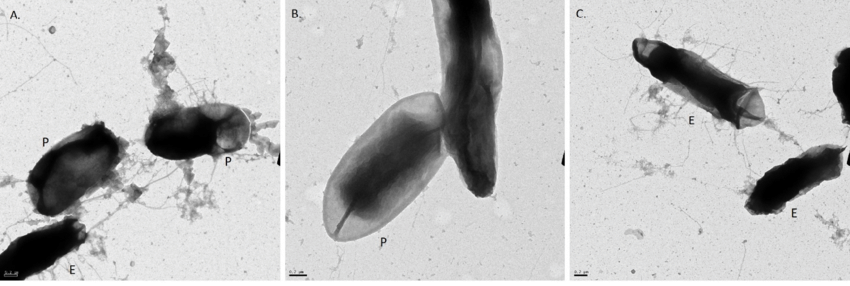
3- La transduction
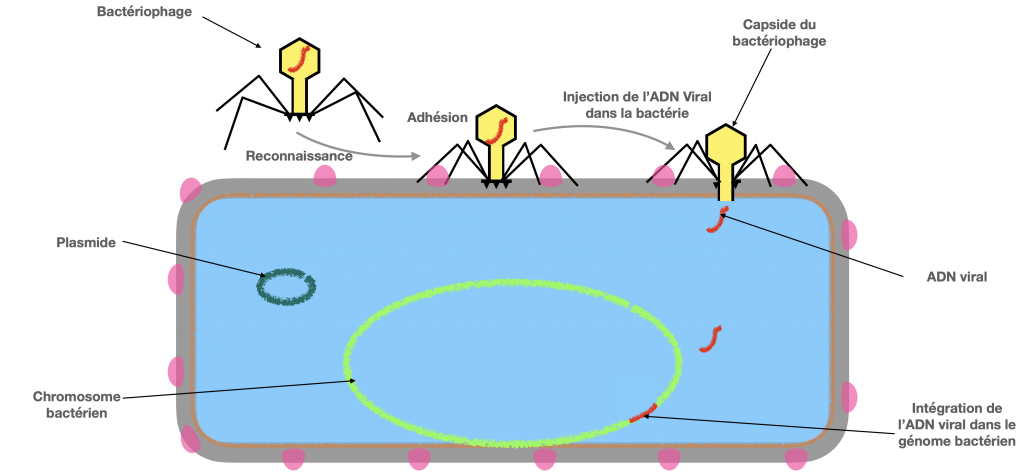
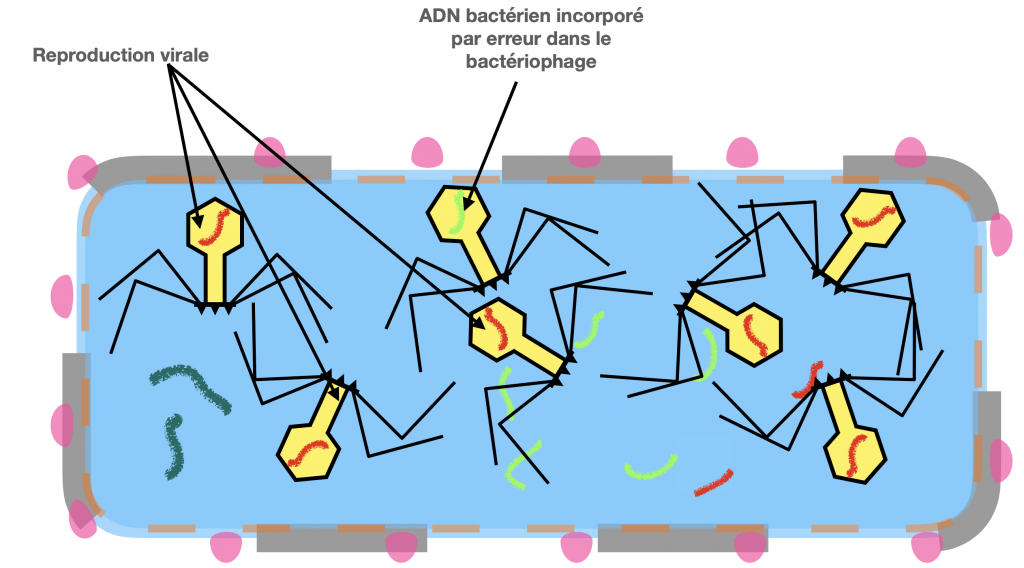
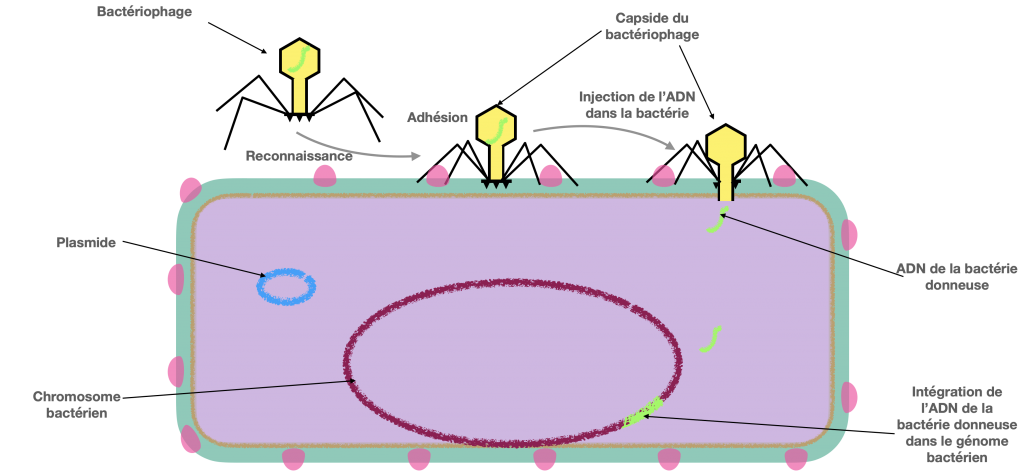
La transduction est un transfert d’ADN bactérien partiel, en utilisant comme vecteur, un virus bactérien qu’on appelle bactériophage et qui n’est plus virulent in fine. Dans la première étape de contamination, il est actif et virulent. Il se fixe à une bactérie donneuse et amorce son cycle de reproduction en insérant sont ADN dans l’ADN bactérien. Puis en associant ses protéines et son ADN, l’un d’entre eux incorpore par erreur de l’ADN bactérien. Ce dernier virus formé va perdre sa virulence, mais pourra tout de même injecter l’ADN qu’il contient dans une bactérie receveuse, donc un ADN bactérien et non viral.
Doc. 7 : Différentes étapes de la transduction.
Doc 7A- Infection par un virus d’une bactérie donneuse
Doc 7B- Prolifération virale, incorporation de matériel génétique de la bactérie dans un virus et destruction de cette dernière
Doc 7C- Intégration par la bactérie receveuse (B) de l’ADN de la bactérie donneuse sans reproduction virale
D- Les transferts horizontaux chez les Eucaryotes
Cette transmission de gènes à d’autres organismes hors descendance n’a que très peu été étudiée chez les eucaryotes : décrit à la fin des années 2000, ce phénomène comporte des caractéristiques (processus, fréquence des transferts) différentes des procaryotes. Chez les Eucaryotes, le transfert horizontal ou hybridation du patrimoine génétique entre deux espèces est plus limité notamment dû au mécanisme de reproduction sexuée. Le taux de transfert peut varier de manière importante entre lignées, et même s’il est relativement faible sur l’ensemble des gènes, il a tout de même lieu. Le plus connu est celui lié aux virus. Chez l’Homme, l’étude de nos gènes montrent que, sur 197 gènes identifiés comme pouvant provenir de transferts horizontaux, leur origine n’est pas liée qu’aux virus.
L’un des exemple couramment cité est celui de la syncitine, protéine d’origine virale, et de la création du placenta chez les mammifères.
Doc. 8 : Origine des gènes hérités par transferts horizontaux chez l’Homme
Un exemple chez les végétaux : https://www.insb.cnrs.fr/fr/cnrsinfo/quand-les-lianes-et-les-arbres-parlent-adn
E- Les transferts horizontaux en santé humaine
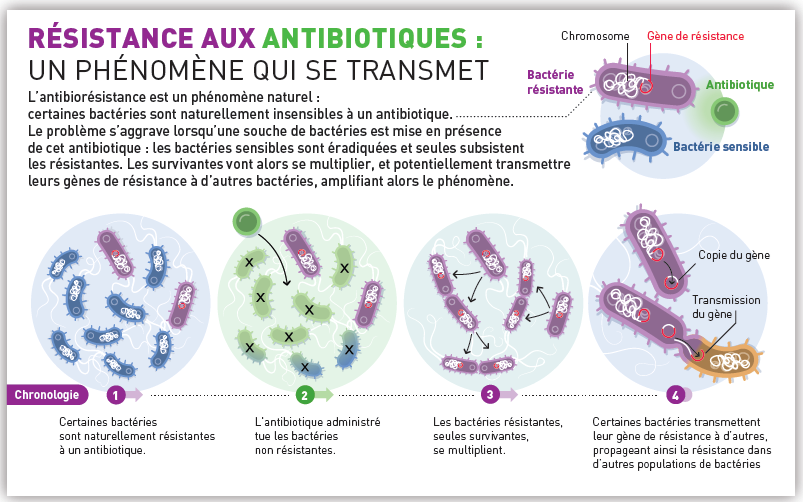
En médecine, les transferts horizontaux sont devenus un enjeu de santé. En effet, des bactéries pathogènes de plus en plus nombreuses acquièrent des propriétés de résistance aux antibiotiques. L’un des exemples les plus célèbres est celui des staphylocoques dorés (staphylococcus aureus) responsables des maladies nosocomiales. Lorsque la barrière cutanée est rompue (blessures), une bactérie cutanée (ou autre) peut alors se développer au niveau de la lésion et provoquer une infection dont le degré de gravité dépendra de la souche bactérienne à l’origine de l’infection ainsi que la rapidité de traitement et l’état de santé général du patient. Mais ce qui est inquiétant, c’est que certaines de ces bactéries ont acquis une résistance à de multiples antibiotiques, notamment en milieu hospitalier. Cette résistance est acquise principalement par le biais de plasmides et donc surtout de phénomène de conjugaison.
Film de l’INSERM – Grandes tueuses : antibiorésistance (13’15‘’)
Doc. 9 : Résistance aux antibiotiques
Notre connaissance des transferts de gènes nous permet aussi de mieux maîtriser la production de médicaments. On parle alors de gènes d’intérêts qui sont transférés dans des cultures bactériennes. On isole les bactéries ayant assimilé le gène d’intérêt, puis elles sont mises en culture. Ces mêmes bactéries produisent alors des molécules ayant un intérêt pharmaceutique pour l’être humain, comme par exemple la production d’Insuline.
II- Les endosymbioses
Définition :
Symbiose : C’est une association durable de 2 organismes tirant bénéfice l’un de l’autre. Exemple : Flore bactérienne de la peau.
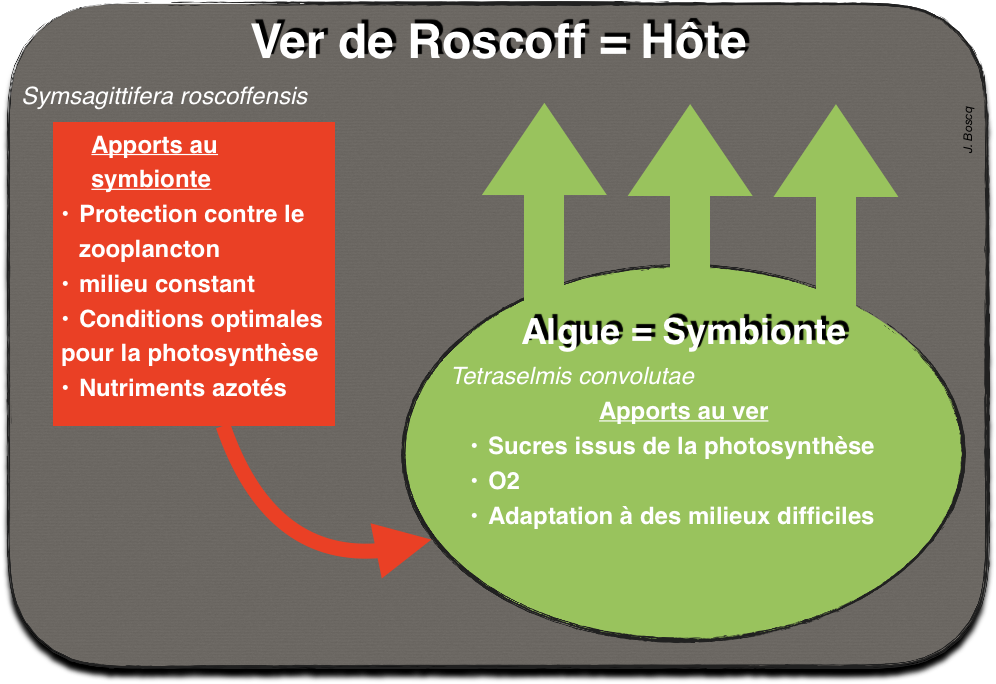
Endosymbiose: Si la symbiose est plus étroite et implique qu’un organisme plus grand, intègre dans ses cellules ou ses organes d‘autres organismes plus petits, on parle alors d’endosymbiose. Exemples : le ver de Roscoff, endomycorhizes…
A- Endosymbiose et diversification du vivant

Endosymbioses
L’endosymbiose est donc une symbiose particulière entre deux organismes vivants, où l’un est contenu par l’autre. L’organisme interne est appelé endosymbionte. Cette association est fréquente dans l’histoire évolutive des eucaryotes. Une fois les organismes fusionnés, la cellule hôte intègre même une part du génome de l’endosymbionte. Parfois ce génome peut aussi régresser au cours des populations successives. Parfois, l’endosymbionte finit même par devenir un organite à part entière de la cellule hôte. Il permet ainsi à l’organisme hôte d’acquérir de nouveaux métabolismes.
Exemple vu en TP des vers de Roscoff
Doc. 10 : Exemples d’échanges entre le ver de Roscoff (Symsagittifera roscoffensis) et son algue Tetraselmis convolutae
B- La théorie endosymbiotique de l’évolution
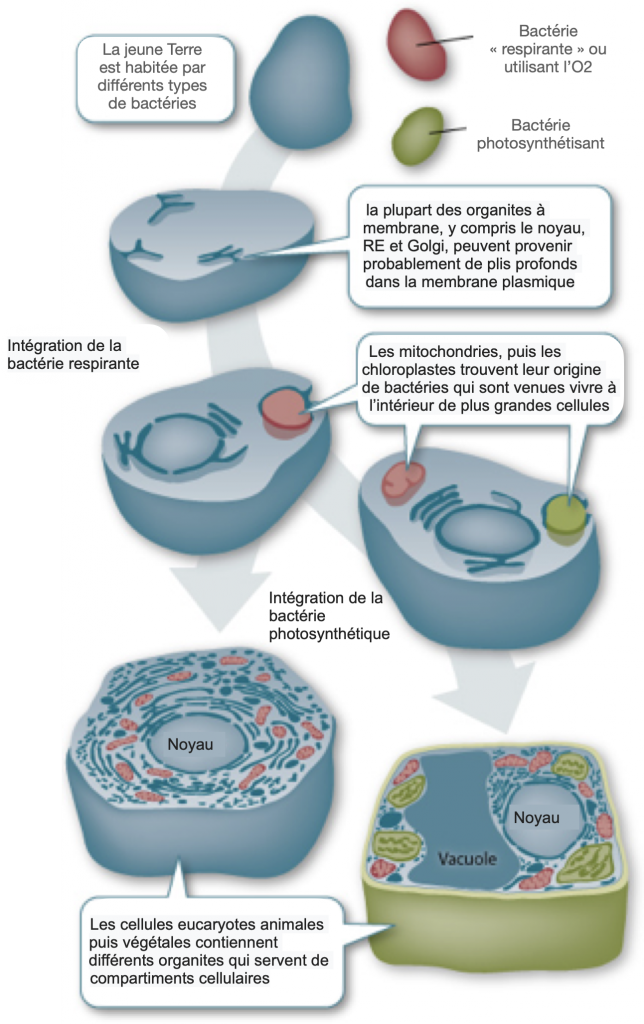
Dans les années 1960, Lynn Margulis proposa une explication endosymbiotique de l’origine de certains organites de la cellule, comme par exemple la mitochondrie ou le chloroplaste. Si cette théorie fut d’abord froidement reçue par la communauté scientifique, elle est aujourd’hui admise et même étendue à d’autres organites.
Ainsi, mitochondries et chloroplastes (ou plus largement les plastes), et même d’autres organites comme les lysosomes ou les flagelles seraient aussi d’anciens procaryotes ayant été incorporé par des cellules plus volumineuses. Ces cellules apportent les nutriments nécessaires aux micro organismes incorporés et ces derniers donnent d’autres capacités à la cellule hôte, lui conférant une meilleure adaptation à son environnement.
Ces grosses cellules anaérobies auraient donc acquis de nouveaux métabolismes tels que la respiration ou la photosynthèse grâce à des procaryotes, qui seraient devenu dans le premier cas les mitochondries et dans le second les chloroplastes des plantes.
Doc. 11 : Théorie endosymbiotique.
C- Des arguments en faveur de la théorie endosymbiotique
L’origine endosymbiotique des mitochondries et des chloroplastes est appuyée par différents arguments :
- Présence d’une double membrane limitant les organites. La membrane externe correspond plus à une membrane plasmique de cellule eucaryote dans sa composition, tandis que la membrane interne correspond plus à une membrane bactérienne.
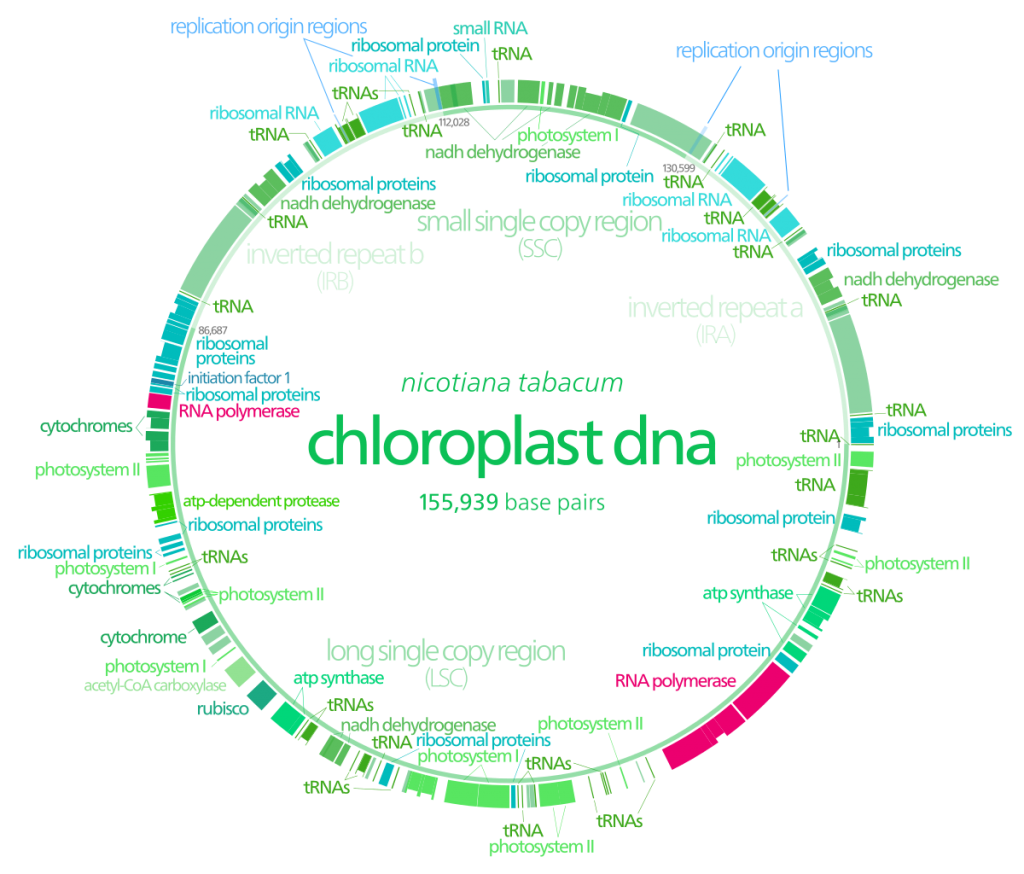
- Les mitochondries et des chloroplastes possèdent en partie leur propre ADN. On dit que ce sont des organites semi-autonomes. Il existe aussi les outils moléculaires permettant la réplication de leurs ADN.
- Il existe des ribosomes particuliers à ces organites que l’on retrouve au niveau bactérien.
- La division des organites intervient à un rythme différent de celui du noyau.
Même si on observe un appauvrissement génétique de ces organites par rapport aux bactéries originelles, on pense qu’une partie du matériel génétique a été incorporé dans l’ADN de la cellule hôte. Cela signifie qu’une partie du génome de l’endosymbionte a été transféré dans le noyau de la cellule hôte ce qui est donc un transfert horizontal.
Doc. 12 : Carte génétique du chloroplaste
{kind=link}
Conclusion :
Le génome de nombreux organismes comporte des gènes qui ont été hérités à l’occasion de transfert entre les différentes espèces. Ces transferts sont dits horizontaux car ils échappent à la logique de la reproduction sexuée et sont transférés d’une espèce à une autre. Pour cette raison on les appelle aussi transferts latéraux. Si les mécanismes chez les procaryotes semblent au nombre de trois (transformation, conjugaison, transduction), chez les eucaryotes les transferts horizontaux sont plus difficiles.
En médecine les transferts horizontaux chez les procaryotes pathogènes compliquent les soins apportés aux patients par l’acquisition d’antibiorésistances multiples, même si en pharmacie on utilise aussi certaines de ces propriétés pour produire des médicaments (gènes d’intérêt).
Les transferts horizontaux ne sont pas les seuls mécanismes de complexification du génome. L’endosymbiose est un autre mécanisme jouant un rôle important dans l’évolution. En effet les mitochondries et des chloroplastes ont trouvé leur place grâce à ces mécanismes. L’ADN que contiennent ces organites en témoigne.
Schéma bilan de votre livre NATHAN :