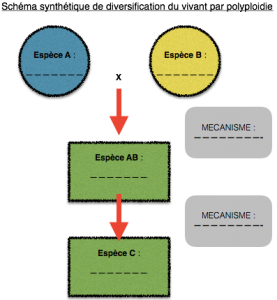
Les mutations ainsi que le brassage génétique associé à la méiose et la fécondation sont des facteurs de diversité au sein d’une espèce. Parfois des accidents de méiose peuvent parfois permettre une évolution, c’est le cas des familles multigéniques. Pourtant, d’autres mécanismes existent, c’est pourquoi au cours de cette séance de TP nous essayerons de découvrir quels autres mécanismes peuvent être à l’origine de la formidable diversité du vivant. Pour chaque diversification, compléter le schéma qui lui correspond le mieux .
I – Tendre ou dur : encore une histoire de chromosomes
(sources d’après http://acces.ens-lyon.fr)
Trois espèces de blé sont cultivées depuis 10.000 ans :
- le blé tendre (Triticum aestivum) espèce dont la culture est actuellement la plus répandue (>90% des surfaces cultivées) ;
- le blé dur (Triticum turgidum=durum)
- l’engrain (Triticum monococcum).
La farine obtenue à partir des grains de blé tendre sert à fabriquer le pain alors que celle obtenue à partir du blé dur est utilisée dans l’industrie des pâtes et des semoules. L’engrain, premier blé cultivé, l’est encore aujourd’hui sur de petites surfaces dans certaines régions montagneuses de Turquie et de Grèce où il sert surtout à l’alimentation du bétail.
Dans certaines régions on trouve encore le Triticum turgidum et le Triticum monococcum à l’état sauvage. En revanche le blé tendre n’a jamais été trouvé à l’état sauvage.
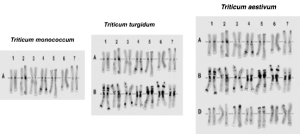
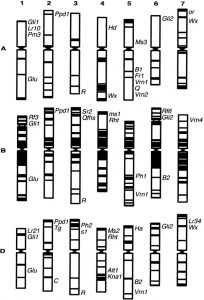
A l’aide des 3 documents fournis et d’anagène© (fichier : Gènes GLU-T.aestivum.edi), expliquez quel lien de parenté il existe entre ces 3 espèces de blés et quel mécanisme pourrait être à l’origine de leur apparition.
Document 1 : Des observations cytologiques ont été réalisées afin de classer les chromosomes des 3 espèces et d’établir les 3 caryotypes suivants :
Document 2 : Carte génétique très simplifiée du génome de Triticum aestivum.
Document 3 : Histoire évolutive d’après les chercheurs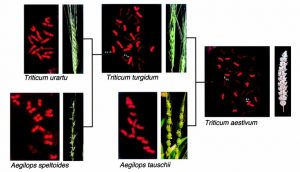
Shéma bilan :
II -Et si les mammifères ne devaient leur existence « qu’à cause » d’un virus ?
A) Échange de gènes entre une microalgue et un virus géant
Pour mieux comprendre, voyons un échange entre une algue et un virus : des échanges possibles : D’après le communiqué de presse du CNRS, expliquez ce qu’est un transfert horizontal de gènes.
d’après : www2.cnrs.fr Paris, 3 août 2009
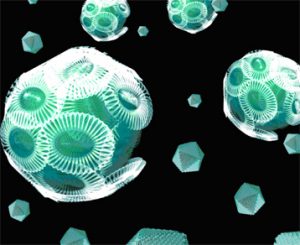 Un transfert de gènes entre la microalgue Emiliania huxleyi et un virus géant vient d’être mis en évidence par des chercheurs du laboratoire Information génomique et structurale (CNRS) et de la Station biologique de Roscoff (CNRS/UMPC), au sein d’une équipe internationale. La particularité de cette découverte est l’implication dans ce transfert de plusieurs enzymes essentielles à toute la voie biosynthétique de la céramide, molécule polyvalente qui participe à divers processus de signalisation cellulaire chez les eucaryotes comme la mortalité cellulaire ou la méiose. Ces travaux sont publiés dans la revue Genome Research d’août 2009.
Un transfert de gènes entre la microalgue Emiliania huxleyi et un virus géant vient d’être mis en évidence par des chercheurs du laboratoire Information génomique et structurale (CNRS) et de la Station biologique de Roscoff (CNRS/UMPC), au sein d’une équipe internationale. La particularité de cette découverte est l’implication dans ce transfert de plusieurs enzymes essentielles à toute la voie biosynthétique de la céramide, molécule polyvalente qui participe à divers processus de signalisation cellulaire chez les eucaryotes comme la mortalité cellulaire ou la méiose. Ces travaux sont publiés dans la revue Genome Research d’août 2009.
Les microorganismes océaniques photosynthétiques (cyanobactéries et microalgues eucaryotes) sont à l’origine de plus de la moitié de la production d’oxygène de la planète. Le niveau élevé de la production biologique de ces microorganismes s’explique par la rapidité de leurs cycles de reproduction et de mort. Il est établi que les virus océaniques jouent un rôle significatif dans ce processus, en infectant et tuant ces microorganismes quotidiennement. Emiliania huxleyi est une microalgue eucaryote à large distribution océanique ; elle est connue pour ses vastes efflorescences et la beauté des écailles de son exosquelette en carbonate de calcium. La mort de ces algues précipite des masses importantes d’exosquelettes dans la lithosphère océanique profonde (ces précipitations sont à l’origine des falaises de craie blanche telles que celles de Douvres, en Angleterre, créée par l’accumulation sur des millions d’années des exosquelettes de ces microalgues).
Un virus géant icosaédrique(1) à ADN nommé EhV est un facteur majeur à l’origine d’une mortalité massive chez E. huxleyi. Dans cet article, les chercheurs ont comparé la séquence génomique de E. huxleyi avec celle du virus EhV. Ils ont ainsi pu établir les preuves d’un transfert de gènes entre cette microalgue eucaryote et le virus. Plus étonnant encore, ce transfert génique implique non pas un seul, mais une série de sept gènes enzymatiques essentiels pour toute la voie biosynthétique de la céramide(2). Le transfert horizontal des gènes est un phénomène bien connu chez les bactéries pathogènes, qui acquièrent par le biais de ce type de transfert de nouvelles fonctions telles que la résistance aux antibiotiques. Cependant, il est rare d’observer le transfert de l’intégralité d’une voie métabolique.
Les auteurs avancent également des preuves du fonctionnement actif de ces gènes de biosynthèse de la céramide tant chez E. huxleyi que les virus. Chez les eucaryotes, la céramide est un élément important des membranes cellulaires, jouant différents rôles dans la réponse au stress et les signaux intracellulaires, y compris le contrôle de l’apoptose(3) et la division cellulaire méiotique. Actuellement, la céramide est utilisée dans certains produits de soin de la peau et anti-âge des industries cosmétiques. Le rôle de cette dernière au sein du virus EhV reste inconnu à ce jour, mais les microbiologistes considèrent que cette nouvelle voie céramide d’origine virale joue un rôle central dans l’interaction entre E. huxleyi et EhV, en contrôlant par exemple la durée de vie des algues hôtes.
B) L’incroyable émergence des mammifères placentaires
Lors de l’implantation de l’embryon dans la paroi de l’utérus, certaines cellules fusionnent entre elles, formant ainsi des cellules « géantes » à plusieurs noyaux qui constitueront le placenta, structure permettant les échanges de nutriments et de dioxygène entre la mère et l’embryon.
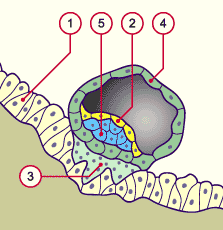 Doc 1 : implantation de l’embryon dans la paroi utérine
Doc 1 : implantation de l’embryon dans la paroi utérine
1- épithélium de la muqueuse utérine
2- hypoblaste
3- syncytiotrophoblaste = futur placenta
4- cytotrophoblaste
5- épiblaste
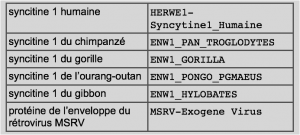
Dans l’ébauche du placenta, les chercheurs ont identifié deux protéines dont la syncytine 1 dont ils pensent qu’elle est impliquée dans la fusion cellulaire. Cette protéine est codée par le gène HERWE1.
Les scientifiques supposent que le gène humain HERWE1 codant pour la syncitine 1 est à l’origine un gène du rétrovirus MSRV (Multiple sclerosis-associated retrovirus).
Afin de tester leur « idée », ils réalisent une expérience de transgénèse (document 1).
Document 2 : Une expérience de transgenèse
On introduit dans des cellules en culture incapables de fusionner entre elles le gène HERWE1.
- a : culture de cellules sans le gène HERWE1
- b : culture de cellules avec le gène HERWE1
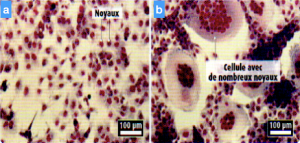
- En quoi les résultats expérimentaux vous permettent-ils de compléter le document 5 si on considère la fusion des cellules permettant la formation du placenta.
- Utiliser les fonctionnalités du logiciel Anagène afin de valider ou invalider cette supposition.
doc. 3 : Cycle des rétrovirus (virus ayant comme matériel génétique de l’ARN)
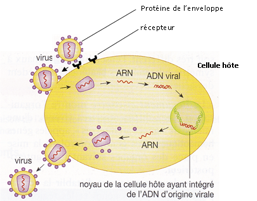
- En utilisant les données du document 3, votre réponse à la question 2 et les données ci-après, justifier l’affirmation « L’Homme est un OGM naturel. »
Données :
Lorsqu’un gène est transféré du génome d’une espèce à celui d’une autre, on parle de transfert horizontal. Lorsqu’un gène est transféré de génération en génération par reproduction sexuée, on parle de transfert vertical.
Schéma bilan :
![]()
III – Etre lézard ou serpent, telle est la question !
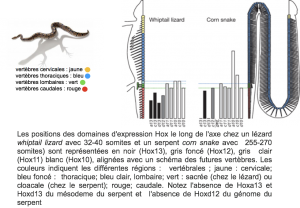
L’équipe du Professeur Duboule de l’Université de Genève a comparé l’expression des gènes Hox chez un serpent et un lézard (Whiptail lizard /red corn snake). Ils ont constaté avec surprise que les segments de génome abritant les gènes Hox étaient bien plus longs chez les squamates que chez les autres reptiles.
«Une accumulation de transposons a vraisemblablement facilité les adaptations qui ont accompagné la transition morphologique entre le modèle ressemblant au lézard et celui du serpent, qui s’est fortement allongé», explique le Professeur.
Une fenêtre d’opportunité s’est présentée au cours de l’évolution du plan de construction des squamates. Il en a résulté un relâchement dans la fonction et la régulation des gènes architectes et un remodelage intégral de la structure des serpents. Ils montrent en particulier des grands changements dans l’expression de Hox13 et Hox10.
Doc. 1 : Gènes homéotiques de Hox 10 à Hox 13 chez le lézard et le serpent.
Définitions :
Somite : Chacun des segments du mésoderme (tissus moyen) de l’embryon, qui donnera notamment les différentes partie du squelette.
Squamates : Ordre de reptiles comprenant les sauriens et les serpents.
Document 2 : Activation du gène Hox6 chez la souris dans tout le corps :
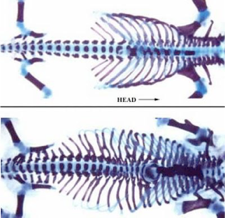 les vertèbres lombaires du mutant portent des côtes comme des vertèbres thoraciques : c’est bien une mutation homéotique. L’existence des côtes sur l’ensemble des vertèbres est un caractère des serpents
les vertèbres lombaires du mutant portent des côtes comme des vertèbres thoraciques : c’est bien une mutation homéotique. L’existence des côtes sur l’ensemble des vertèbres est un caractère des serpents
1) D’après ces documents, dites ce que sont les gènes Hox
2) Quels changements sont décrits dans le document 7 concernant les gènes Hox 10 et 13 ?
3) Qu’apporte le document 8 concernant le gène Hox 6 ?
Schéma bilan :