
Cours : L’inéluctable évolution des génomes au sein des populations
Mise à jour le 8/04/2025 par J. Boscq
Introduction :
Les études génétiques par croisements simples ou croisements tests permettent de montrer ou de définir les différentes possibilités de descendance d’individus à partir d’un couple. Si certaines mutations, ou même des transferts de gènes sont avantageux en terme d’adaptation à l’environnement, alors en toute logique, ces nouveaux gènes doivent persister dans une population. La génétique des populations est une science qui analyse l’ampleur et la distribution de la variation génétique au sein des populations ainsi que les forces évolutives qui contrôlent cette variation.
La biodiversité change, elle évolue sans cesse grâce à l’évolution des génomes, mais également en fonction de son interaction avec l’environnement.
Photo de couverture : d’après Jboscq
Problèmes : Comment expliquer la stabilité ou l’évolution d’une population, d’une espèce ? Est-il possible de modéliser l’évolution des fréquences d’allèles au sein d’une population ?
I- L’équilibre de Hardy-Weinberg et ses limites
A- Stabilité de fréquence d’allèles dans une population

Définitions :
Population : ensemble d’individus d’une même espèce regroupés ensemble (en un même moment et un même lieu) pouvant interagir entre eux et se reproduire.
Espèce : Ensemble d’individus, vivants ou fossiles, à la fois semblables par leurs formes adultes et embryonnaires et par leur génotype, vivant au contact les uns des autres, se reproduisant entre eux et dont la descendance est également féconde. Une espèce est composée de plusieurs populations.
Panmixie : Reproduction d’individus par des unions faites au hasard, en l’absence de sélection naturelle.
Pool génétique : c‘est la somme, à un moment donné, de tous les allèles des membres d’une population susceptibles d’avoir des descendants (voir doc 1).
1- Préalable : Fréquences de génotypes et fréquence d’allèles
Dans une population donnée, et quand on étudie un gène, il existe un certain nombre de génotype et d’allèles. Les deux notions ne sont pas à confondre. Pour mieux comprendre, nous allons utiliser l’exemple de lapins :
Doc 1 : Représentation d’une population de lapins dans leur milieu à un instant t

Analyse du document 1 :
On peut déterminer 2 allèles A et a
- Les homozygotes AA et hétérozygotes Aa sont les génotypes responsables du phénotype [poils Gris] et les homozygotes aa sont les génotypes responsables du phénotype [poils jaunes]
- La proportion des phénotypes est presque de 1/4 de lapins aux [poils jaunes] (5/21 x 100 = 23,8 %) et 3/4 de lapins aux [poils Gris] (16/21 x100 = 76,2 %)
- Les effectifs en fonction des génotypes sont : AA = 6 ; Aa = 10 ; aa = 5
- Mais le nombre d’allèles est le suivant dans la population: A= (2 x 6 (AA) + 10 (Aa)) = 22 et a = (2 x 5 (aa) + 10 (Aa)) = 20 et donc la proportion des allèles est :
- A : 22/42 x 100 = 52,4 %
- a : 20/42 x 100 = 47,6 %
Ainsi on peut étudier la fréquence allélique de la population :
La fréquence de A est alors appelée p et la fréquence de a est appelée q.
On s’aperçoit alors que p + q = 1. (réciproques : p = 1 – q ou q = 1 – p )
2- La loi de Hardy-Weinberg
La stabilité des fréquences alléliques d’une population ou équilibre a été étudiée indépendamment par le mathématicien anglais Hardy et le médecin allemand Weinberg au début du XXème siècle. Cela a permis d’aboutir à un modèle mathématique d’équilibre génétique des populations, appelée loi de Hardy-Weinberg. Cette dernière postule qu’au sein d’une population, il y a un équilibre des fréquences alléliques et génotypiques d’une génération à l’autre, ce qui signifie que ces fréquences restent constantes dans la population au cours des générations successives.
Reprenons nos fréquences d’allèles précédemment citées :
avec p la fréquence de l’allèle A et q la fréquence de l’allèle a ; Rappel : p+q =1
La fréquence d’un allèle dans le pool génétique est égal à la probabilité que cet allèle soit choisi lorsqu’on prélève au hasard un allèle du pool génétique pour former un ovule ou un spermatozoïde.
Concernant nos lapins :
On peut calculer la probabilité qu’un lapin de la génération suivante soit homozygote, soit A//A. Pour prélever le premier A, la probabilité est de 0,524 (52,4%) et autant pour le 2e A. Le produit de ces deux probabilités, [(0,524)2 = 0,274576] correspond à p2.
On peut de la même manière calculer la probabilité qu’un lapin de la génération suivante soit homozygote a//a et on aboutit à q2 [soit (0,476)2 = 0,226576].
Enfin pour calculer la probabilité d’être hétérozygote (A//a), on peut d’abord prélever 1A avec la probabilité P, puis 1a avec la probabilité q : soit p x q. Mais on peut aussi prélever un a en premier puis un A ensuite soitq x p. Donc la probabilité qu’un lapin de la génération suivante soit hétérozygote A//a est de pq + qp soit 2 pq [ 2 x (0,524 x 0,476) = 0,498848].
On remarque d’ailleurs que la somme des probabilités fait aussi 1 : 0,274576 + 226576 + 0,498848 =1
On peut alors généraliser cette loi à l’ensemble des populations sous cette forme :
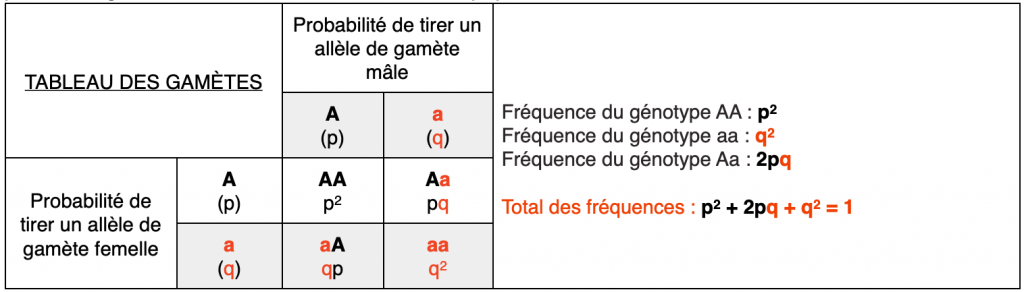
Enfin on peut alors revenir sur la fréquence de l’allèle A f(A) et sur la fréquence de l’allèle a f(a) :
- f(A) = p2 + pq = p (p + q) = p
- f(a) = q2 + pq = q (q + p) = q
Car p + q = 1
Les conditions pour atteindre l’équilibre de Hardy-Weinberg sont nombreuses, et ne sont donc que rarement remplies. En effet, le vivant est en constante évolution… Néanmoins, cet équilibre permet de voir si la population évolue ou non.
B-Limite de la loi et mise en application

Loi de Hardy Weinberg
La loi de Hardy-Weinberg est soumise à conditions :
- La population étudiée doit être de grand effectif
- Les unions se font au hasard (= panmixie) – il n’y a pas de préférences sexuelles.
- Il n’existe ni migration, ni sélection naturelle,
- Le taux de mutations est constant
- Les proportions des différents génotypes restent constantes d’une génération à l’autre (pas de flux entrants tels que les transferts de gènes ou autres)
Cela permet d’imaginer une population sans évolution… Si cet équilibre n’est pas respecté, alors la population est entrain d’évoluer et il faut analyser pourquoi.
Considérons les fréquences du modèle de Hardy Weinberg : fmHW
Et considérons les fréquences réellement observées : fr
- Si la population montre une fréquence pour le gène étudié fr = fmHW, alors elle est à l’équilibre de Hardy-Weinberg.
- Si la population présente une fréquence pour le gène étudié fr ≠ fmHW, alors la population est instable.
Dans ce dernier cas, la population étudiée ne respecte pas l’une des conditions ci-dessus, elle continue d’évoluer pour ce gène.
II- Les forces évolutives

Battle de QCM
Les cinq conditions du principe de Hardy-Weinberg indiquent donc, si elles varient, des possibles changements évolutifs des populations. En fait, que ce soit des mutations, des variations de flux de gènes, de l‘accouplement non aléatoire, de la dérive génétique dans de petites populations ou des pressions de sélection naturelle, chacun de ses événements peut entraîner des changements dans les proportions d’allèles ou de génotypes.
Définition :
Spéciation : Formation d’espèces nouvelles. (Il y a spéciation lorsque des mutants ne peuvent s’hybrider avec les formes anciennes [amixie].)
A- Les mutations changent les allèles
Même si le taux est faible, la mutation est la cause ultime de variation génétique et permet l’évolution. Un gène type mute environ une fois sur 100 000 divisions cellulaires. Les mutations sont aléatoires et n’apparaissent pas plus en cas de contrainte sélective du milieu. Par contre elles subsistent dans la population, si elles touchent les cellules germinales, que ce soit des mutations à caractère dominant ou à caractère récessif.
Doc.2 : mutations et sélection naturelle
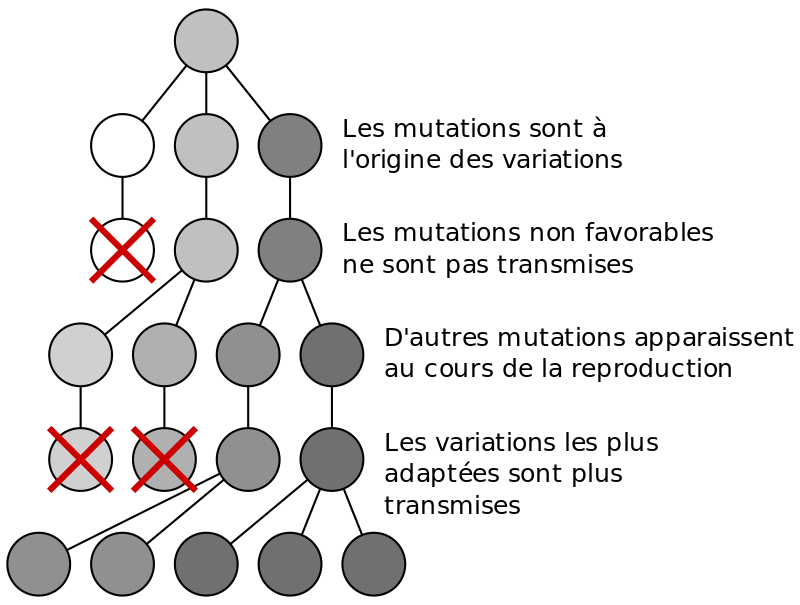
{kind=link}
D’après https://planet-terre.ens-lyon.fr
{kind=link}
B- Flux de gènes
Le flux génique est le passage d’allèles d’une population à une autre. Cela peut être un puissant facteur de changement. Parfois il est évident, car on observe un déplacement d’un individu dans une autre population. Si cet individu présente une adaptation plus appropriée ou une préférence sexuelle alors la population d’accueil pourra être modifiée. Parfois le flux génique est beaucoup plus subtile, comme par exemple le transport de gamètes chez les plantes d’une population à une autre. Certains animaux sont capables de transporter des pollens (zoogamie) sur des longues distances. C’est aussi le cas pour le transport des graines (zoochorie).
Doc.3A : Pollinisation d’une tulipe, population appartenant à une culture d’un champ. Mais dans le champ voisin, des tulipes d’une autre couleur sont cultivées

Doc.3B : modélisation du flux de gènes chez les plantes.

Complément :
Le flux de gènes présente donc un risque potentiel associé aux grandes cultures d’OGM. … des champs de culture non GM doivent côtoyer des champs de culture GM ; Il y a donc des risques de croisements entre une plante transgénique et une variété non transgénique de cette plante (croisements intervariétaux) ou entre une plante transgénique et une espèce sauvage apparentée (croisements interspécifiques donnant des hybrides interspécifiques). Donc ces risques de flux de gènes – c’est-à-dire de transferts de la résistance de la plante transgénique et de dissémination de cette résistance dans l’environnement – peut créer de nouvelles super espèces, si celles-ci ont un avantage sélectif.
C- Sélection sexuelle ou accouplements non aléatoires
Les individus ayant un certain génotype s’accouplent parfois avec un autre plus souvent que prévu par le hasard, un phénomène connu sous le nom d’accouplement non aléatoire. On parle de préférence sexuelle.
Par exemple, l’homogamie définit l’accouplement d’individus phénotypiquement semblables, c’est donc un type de reproduction non aléatoire. Cela explique que les fréquences de génotypes particuliers diffèrent grandement de ceux prévus par le principe d’Hardy-Weinberg.
Un autre exemple est à prendre chez les vertébrés, car beaucoup d’espèces favorise la préférence sexuelle qui ira à un mâle dominant. Ce dernier est défini par sa force, sa vigueur et donc sa capacité à mieux survivre dans son environnement (cela rejoint le principe de sélection naturelle). Il ne s’agit donc pas de panmixie.

Complément :
Chez les plantes, les autogames stricts ne permettent pas non plus une panmixie.
Un autre exemple très proche est l’endogamie : c’est la conséquence d’un accouplement entre individus apparentés. Puisque ceux-ci sont génétiquement semblables, l’endogamie aura le même résultat que l’homogamie et augmentera la proportion d’homozygotes dans la nature.
La panmixie, quant à elle, favorise un entretien des hétérozygotes.
D- La dérive génétique modifie les fréquences alléliques
Au sein de petites populations, le hasard peut modifier fortement les fréquences des allèles. De tels changements aléatoires de ses fréquences donnent l’impression que les fréquences dérivent d’où l’expression : dérive génétique.

{kind=link}
C’est pour cette raison qu’une population doit avoir une grande taille (au dessus d’un seuil) pour atteindre l’équilibre de Hardy-Weinberg. Sinon la dérive génétique peut aboutir à la perte d’allèles dans des populations isolées. Les allèles qui initialement sont les plus rares sont particulièrement vulnérables.
1- cas particulier de l’effet fondateur
Parfois ou un ou quelques individus migrent et fondent une nouvelle population qui s’isole. Il est probable que ces fondateurs ne possèdent pas tous les allèles de la population originelle dont ils sont issus. Cela signifie que certains allèles peuvent être définitivement perdus. Mais à l’inverse, certains allèles rares peuvent tout à coup devenir une part génétique fondamentale de la nouvelle population. C’est ce que l’on appelle l’effet fondateur.
Doc.4 : L’effet fondateur
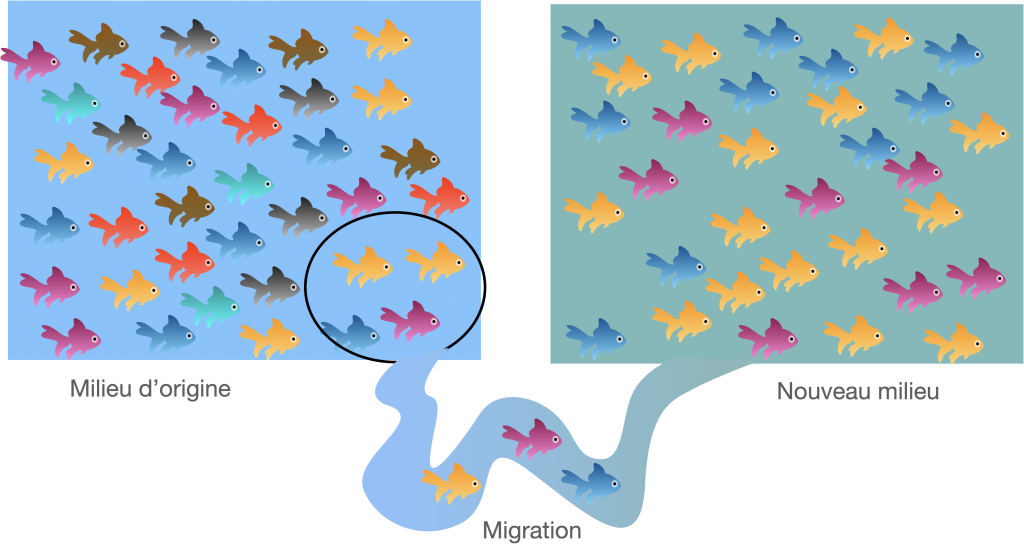
2- cas particulier de l’effet goulot d’étranglement
Certaines populations peuvent se voir réduire de taille drastiquement à cause de différents événements. En effet, une catastrophe naturelle, une inondation, ou même une épidémie peuvent être à l’origine d’une mortalité subite de la presque totalité des individus. Les quelques survivants constituent à leur tour une population d’origine. Mais, cela aboutit à un appauvrissement allélique du groupe. Le manque de variabilité est un risque à l’extinction.
Doc.5 : L’effet goulot d’étranglement
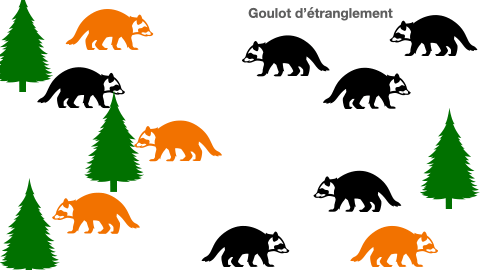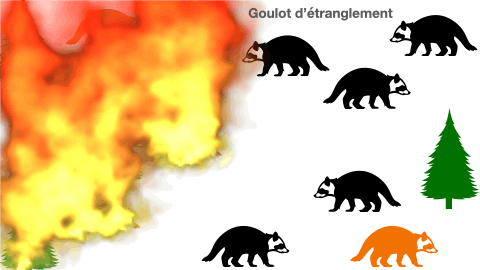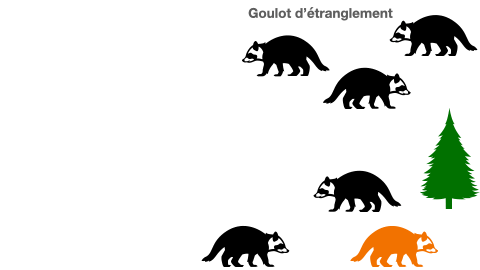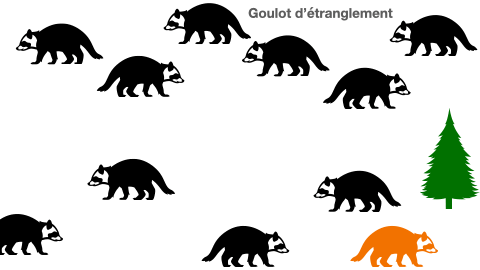
E- Sélection naturelle
Ce processus participant à l’évolution a été largement décrit par Charles Darwin lorsqu’il publie en 1859 « De l’origine des espèces », et est toujours d’actualité aujourd’hui. On peut décrire la sélection naturelle comme suit : La variabilité génétique offre à chaque génération de nouveaux phénotypes, seuls ceux qui procurent un avantage adaptatif même minime sont retenus par la sélection naturelle, qui se révèle stabilisante. En revanche, lors des radiations adaptatives, elle favorise l’exploitation de nouvelles ressources, la sélection naturelle est alors diversifiante. Dans les deux cas, la sélection naturelle œuvre pour une adaptation la meilleure des organismes à leur milieu. La sélection dépend donc de la pression de l’environnement et de la capacité d’adaptation des individus. On peut observer différentes sélections parmi lesquelles :
- Une sélection pour éviter les prédateurs.
- Une sélection pour s’adapter aux conditions climatiques, ou au changements topographiques
- Une sélection pour s’adapter à la pression exercée par l’homme …/…
Doc.6 :La sélection naturelle observée par Darwin chez les pinsons des Galapagos –
Doc.6 A: les pinsons de Darwin

Doc.6 B : L’évènement de sélection de 1977

▪ La taille du bec étant un caractère héréditaire, leur descendance a un gros bec
Doc.6 C : Compétition et sélection
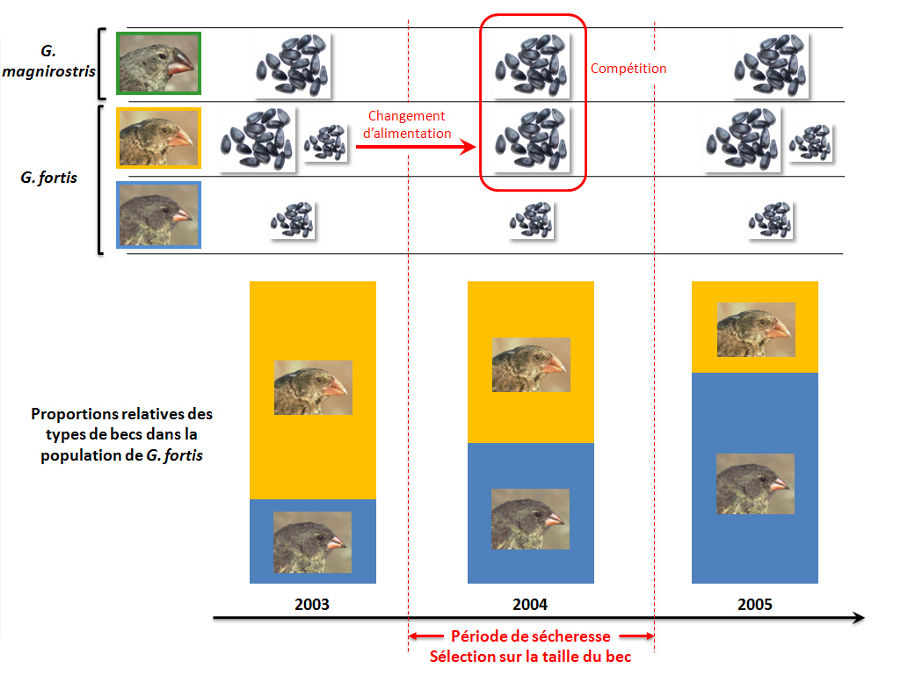
Ainsi, dans les écosystèmes, les facteurs ne sont pas stable au cours du temps. Et c’est le cas des facteurs abiotiques (physico chimiques) et des facteurs biotiques (compétition, prédation, reproduction…). Les peuplement peuvent alors changer. Les individus qui pouvaient être adaptés à un milieu ne le sont pas forcément au milieu suivant. La population la plus adaptée s’installe, l’évolution est en marche.
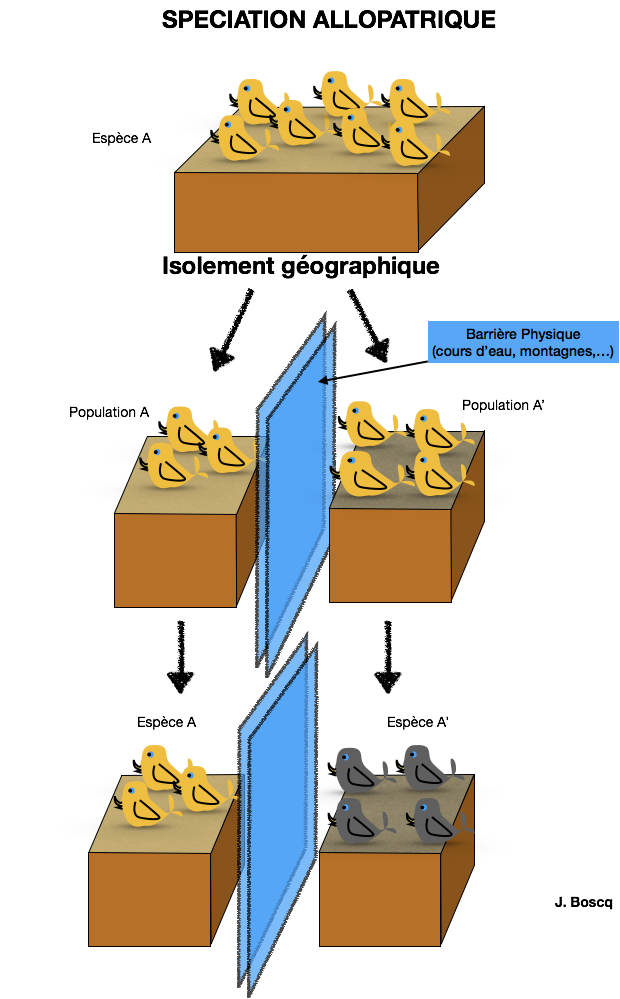
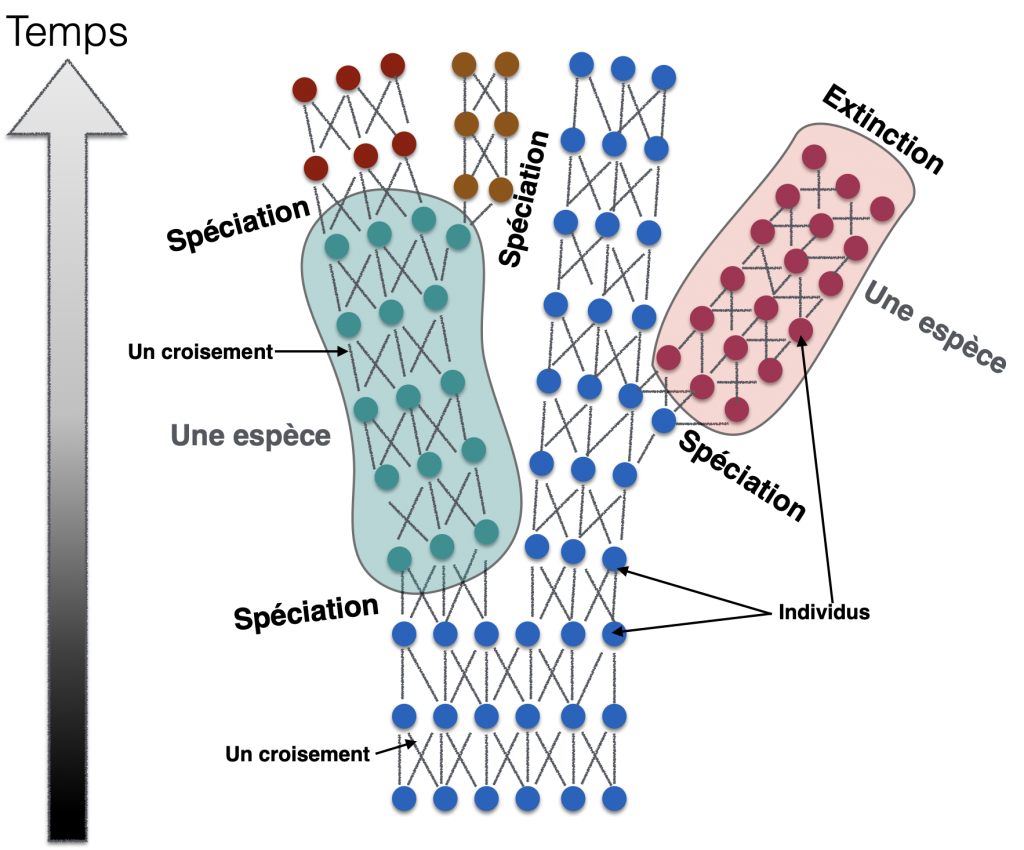
III- Spéciation ou extinction
Au regard des populations, on peut considérer une espèce comme une population d’individus suffisamment isolés génétiquement d’autres populations. Ainsi, elle n’existe que pendant un temps limité. Si les individus de cette population disparaissent, le plus souvent pour des raisons de modifications de milieu, on parle d’extinction. Mais si un nouvel ensemble d’individus semble se singulariser, alors on parle de spéciation. Ce qui caractérisera définitivement la spéciation est l’isolement reproductif. On distingue entre autre 2 types de spéciation en fonction de cet isolement :
- ALLOPATRISME : Si l’isolement est géographique (modification du relief, du climat, ou du milieu), 2 populations vont évoluer indépendamment jusqu’à ce que la dérive génétique imprime des différences telles que si les 2 populations sont replacées ensemble, elles ne sont plus interfécondes.
- SYMPATRISME : S’il n’y a pas d’isolement géographique, la sélection naturelle va favoriser des individus plus adaptés au milieu et donc leur reproduction entre eux. Il pourra donc y avoir coexistence de 2 populations qui si l’isolement reproductif est atteint, aboutit à deux espèces distinctes.
Doc.7 : Allopatrisme et sympatrisme
IV- La notion d’espèce
La biodiversité est la diversité du vivant, c’est-à-dire la diversité des espèces. Appartiennent à une même espèce des individus qui se ressemblent, qui possèdent le même stock chromosomique, qui peuvent se reproduire ensemble et dont la descendance est féconde. Au cours de l’histoire, la notion d’espèce s’est enrichie des découvertes, dont celles de la génétique et de l’évolution. Ainsi les critères phénotypiques et d’inter-fécondité font partie intégrante de cette définition. La notion de flux de gènes s’est ajoutée ainsi que les comparaisons moléculaires permettant de définir une parenté.
Doc.8 : L’ADN barcoding ou la détection d’espèces dans un milieu.
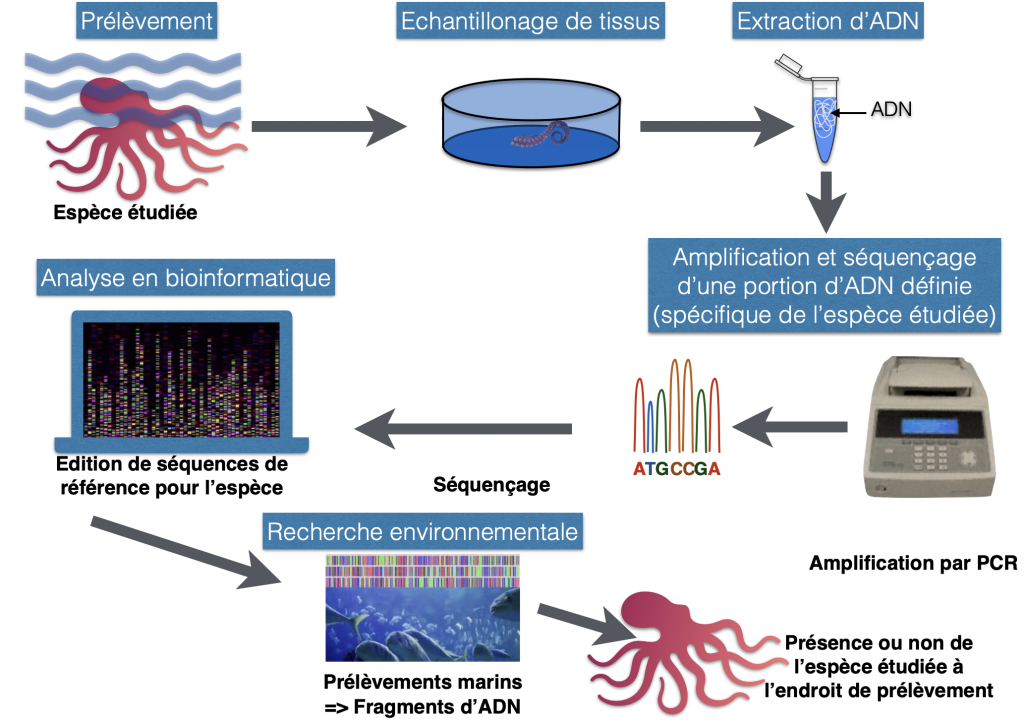
Complément : l’ADN barcoding ou la recherche d’espèces dans la nature
Un barcode moléculaire est un fragment d’ADN présent chez tous les organismes vivants. La séquence de ce fragment d’ADN est quasiment identique chez des individus qui appartiennent à la même espèce, et permet donc de déterminer l’espèce à laquelle appartient un individu en ne connaissant que la séquence de ce fragment d’ADN. Le fragment choisi est un gène du génome mitochondrial codant pour une protéine qui intervient dans la chaîne respiratoire de la mitochondrie. Chaque cellule contenant de nombreuses mitochondries, le gène COI est présent en de nombreuses copies, ce qui facilite son séquençage. De plus, ce gène présente un niveau de variabilité intéressant : les différences entre les séquences de ce gène chez différents individus, apparues par mutations au cours du temps, sont faibles entre les individus d’une même espèce et élevées entre des individus d’espèces différentes.

C’est ainsi que cette séquence d’ADN peut être utilisée, à l’instar des code-barres utilisés dans le commerce, comme un outil d’identification taxonomique des organismes vivants : tous les individus d’une espèce ont des séquences identiques ou très proches, ce qui va permettre l’identification d’un spécimen en comparant sa séquence avec celles des espèces connues.
D’après planetvie.ens.fr
Conclusion : Schéma bilan
Pour aller plus loin : Explication du barcoding